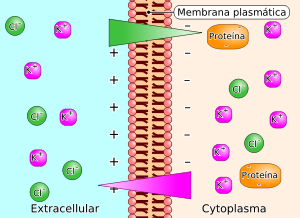
El agua es considerada como la molécula más importante de la vida, y la capacidad de un organismo para hacer frente a los cambios en su contenido de agua interna es esencial para la supervivencia. En particular, la pérdida de agua interna es una amenaza común, la evaporación en el aire, durante la excreción de desechos o de ósmosis en un entorno acuoso concentrado. Este último puede ocurrir en un ambiente salino externo, vía extracelular, o por enfermedades que causan desequilibrios osmóticos (por ejemplo, la diabetes y sus asociados).
Tradicionalmente, organismos se han dividido en dos grandes categorías en términos de las adaptaciones al estrés hídrico: osmoconformadores, que suelen utilizar osmolitos orgánicos para mantener la presión osmótica celular igual a la del medio ambiente externo, y osmorreguladores que utilizan el transporte de iones homeostáticamente para regular la presión osmótica interna.
Los osmoconformadores se encuentran más comúnmente en los océanos e incluyen la mayoría de las otras formas de vida que la mayoría de los vertebrados y algunos artrópodos. Las sales (principalmente NaCl) de agua de los océanos tienen una concentración osmótica del medio de ~ 1000 · miliosmoles por litro (1000 · mOsm), muy por encima del ~ 300 a 400 mOsm · creados por los solutos básicos que se encuentran en la mayoría de las células (K, metabolitos, proteínas, etc.) Para evitar la contracción osmótica, los fluidos internos de osmoconformadores marinos sobre la misma presión osmótica que su entorno (por ejemplo, 1000 mOsm ·).
Estos animales marinos suelen tener órganos reguladores (por ejemplo, las branquias, los riñones) que trabajan para mantener los fluidos internos del cuerpo a ~ 400 mOsm. Este es el patrón heredado por los vertebrados terrestres, que suelen tener ~ 300 · fluidos corporales mOsm.
Los solutos orgánicos similares o idénticos a los osmolitos orgánicos también son acumulados por algunos organismos bajo estrés térmico y anhidrobiótico y posiblemente bajo presión hidrostática. Estos solutos son generalmente llamados 'solutos compatibles', basado en el concepto de no perturbar las macromoléculas celulares, incluso cuando los solutos se encuentran en altas concentraciones.
TIPOS DE OSMOLITOS ORGANICOS
Se sabe que muchas pequeñas moléculas diferentes sirven como osmolitos orgánicos y otros solutos compatibles. Estos solutos caen en una categoría química como son: los hidratos de carbono pequeños, incluyendo los azúcares (trehalosa, por ejemplo), polioles (glicerol, inositoles, sorbitol, etc) y derivados (por ejemplo, como o-metil-inositol), aminoácidos (glicina, prolina, taurina, etc) y derivados (por ejemplo, ectoine); metilaminas [como N-óxido de trimetilamina (OTMA) y la glicina betaína] y solutos de metilsulfonio incluyendo dimetilsulfonopropiato (DMSP), y la urea.
PROTECCION METABOLICA
La taurina es uno de las principales, osmolitos dominantes en muchos invertebrados marinos como bivalvos en aguas poco profundas. La taurina también es relativamente alta en el corazón de los mamíferos y en las células del cerebro, donde puede servir como osmolito importante para evitar casos graves de deshidratación.
También es esencial para el desarrollo neuronal de los mamíferos de manera que no puede estar relacionado con el equilibrio osmótico. Este compuesto citoprotector actúa como un antioxidante, un modulador de calcio, un neuromodulador sináptico y un estabilizador de la membrana.
ANTIOXIDACION
En algunos casos, osmolitos pueden ser compatibles y al mismo tiempo citoprotectores activos que actúan como antioxidantes. Por ejemplo, se ha encontrado que los ciclitoles (polioles cíclicos) y polioles como el manitol, que son utilizados por muchas plantas para retener agua, también puede eliminar los radicales libres generados durante la sequía y el frío y otros factores de estrés.
De todos los solutos acumulados en concentraciones relativamente altas en algunas situaciones, la hipotaurina, con su átomo de azufre reactivo, es uno de los más fuertes antioxidantes, capaces de recoger radicales OH (convirtiendo la hipotaurina en taurina), así como HOCl.
BALANCE REDOX Y PROTECCION DE HIPOXIA
El glicerol, el arquetípico soluto compatible, se acumula en algunas levaduras y algas. Se ha demostrado que es compatible en gra medida con la función de las proteínas, pero su síntesis también requiere el uso de NADH. Esto es esencial para mantener el equilibrio redox celular (por la regeneración de NAD) durante el metabolismo anaeróbico, de hecho, levaduras mutantes incapaces de hacer que el glicerol no sólo son altamente sensibles al estrés osmótico, pero también se acumula NADH excesivo y por lo tanto no puede crecer. El Glicerol también ayuda a reducir la producción de radicales de oxígeno. La acumulación de prolina como osmolito en plantas que sufren estrés hídrico mantiene los estados redox, para estabilizar las propiedades osmolares. Otros osmolitos pueden proteger a las células durante la hipoxia por otros mecanismos. La betaína β-alanina, un osmolito importante en varias especies de plantas de marisma, parece sustituir a la glicina betaína que se encuentra en las plantas afines. A diferencia de glicina betaína, β-alanina, la betaína no requiere el uso directo de oxígeno para su producción, posiblemente favoreciendo su uso en condiciones de hipoxia.
OTRAS FUNCIONES METABOLICAS Y LA COMPATIBILIDAD REVISADA
Otras importantes funciones metabólicas y de protección se atribuyen a algunos osmolitos. Los carbohidratos como osmolitos como la glucosa, sorbitol y trehalosa puede servir como fuente inmediata de energía después de que un organismo emerge de un letargo inducido por el estrés. La defensa contra los depredadores es otra posible función de algunos osmolitos.
ESTABILIZACION Y OPOSICION
En la naturaleza, la estabilización de la capacidad parece ser usado sólo cuando hay estrés que directamente desestabiliza macromoléculas y membranas. Este estrés incluye perturbación de solutos, anhidrobiosis, alta
temperatura, congelación y alta presión hidrostática.
PERTURBACION DE SOLUTOS: UREA Y SALES
Algunos osmolitos orgánicos son capaces de compensar, o "contrarrestar", los efectos de los solutos que también se acumulan en estrés osmótico y que perturban macromoléculas. La urea es un perturbador. Se trata de residuos altamente concentrados que producen los riñones de mamíferos. La urea desestabiliza muchas estructuras macromoleculares e inhibe funciones tales como las uniones ligando. La neutralización de la urea ha sido ampliamente confirmada en una variedad de sistemas de proteínas y también se ha demostrado recientemente para ácidos nucleicos en forma de tRNA de bacterias.
Las metilaminas también puede compensar algunos efectos perturbadores de las sales. Los derivados metilados de la glicina (sarcosina, dimetilglicina y betaína glicina) puede contrarrestar la inhibición de la actividad de NaCl de una enzima de la planta, la protección aumenta con el grado de metilación.
Muchos otros estudios muestran neutralización de la inhibición de la sal por metilaminas, incluyendo los complejos sistemas celulares.
ANHIDROBIOSIS
Los disacáridos, especialmente la trehalosa, comúnmente es un anhidrobiótico en organismos latentes. Sin embargo, estos azúcares no presentan compatibilidad no interactiva y no son osmolitos ya que los organismos pierden la mayor parte de su agua. Más bien, estos solutos parece que se unen a las macromoléculas y membranas, en esencia para reemplazar las moléculas de agua y mantener la estructura básica de estas biomoléculas de gran tamaño. Por otra parte, la trehalosa forma un estado similar al vidrio (es decir, vitrifica) en estado seco, que también ayuda a preservar las estructuras celulares.
CONGELACION
La congelación es otro tipo de estrés que enfrentan muchos ectotermos. Las estrategias para sobrevivir a temperaturas corporales por debajo del punto de congelación se divide en dos categorías:
Evasión de la congelación y tolerancia al hielo. Los evasores (en cuyo cuerpo los líquidos no se congelan) utilizan una variedad de mecanismos tales como proteínas anticongelantes desconocidas, sitios de nucleación y sobreenfriamiento reducido.
Muchos evasores también acumulan altos niveles de anticongelantes o crioprotectores, que suelen ser compatibles con los hidratos de carbono tales como el glicerol.
Los carbohidratos también se encuentran como crioprotectores en muchas plantas.
Por lo tanto, los hidratos de carbono pequeños han sido seleccionados como anticongelantes coligativos de forma independiente en diferentes taxones y estrategias. Además, ciertos aminoácidos como la prolina también se acumulan en algunos animales que toleran el frío. De hecho, hay evidencia de que los crioprotectores pueden clasificarse en dos categorías con distintas funciones.
En primer lugar, los hidratos de carbono, tales como glicerol actúa como anticongelantes, y, en la tolerancia al congelamiento, como osmolitos (es decir, que reducen la pérdida de agua celular), mientras que al mismo tiempo es compatible con macromoléculas. Por el contrario, un segundo grupo de crioprotectores pueden tener la estabilización de las funciones que otros solutos no. En particular, la prolina y la trehalosa parece que se unen a la cabeza de los grupos de fosfolípidos de la membrana, sustituyendo las moléculas de agua. Por lo tanto, pueden estabilizar las membranas durante la contracción de la célula.
ALTAS TEMPERATURAS
Casi todos los osmolitos naturales y otros solutos compatibles pueden aumentar la estabilidad térmica de la proteína in vitro, aunque para la mayoría de osmolitos, esto ocurre sólo en concentraciones altas no fisiológicas. Sin embargo, ciertos solutos de hidratos de carbono puede ser utilizados en los organismos vivos para contrarrestar la interrupción de la temperatura de las proteínas. Por ejemplo, el estrés por calor induce la acumulación de trehalosa en levaduras, en los que el disacárido puede proteger las enzimas de la desnaturalización térmica.